Qu'est-ce que le Paludisme
A. Population exposée et répartition géographique :II. Plasmodium : un parasite adapté à son hôte.
B. Mortalité due au paludisme :
C. Une situation alarmante :
Les
familles polygéniques sont un mécanisme intervenant dans
une telle dynamique, générant une diversité accrue
et permettant une adaptation rapide. Nous avons étudié, au
cours de ce travail, deux gènes de la sous-famille Pf60, appartenant
à la super famille Pf60/var de Plasmodium falciparum,
dans lespoir de découvrir leur fonction, de préciser leurs
relations avec la sous-famille var et dintégrer ces dernières
dans un schéma plus général de génération
de diversité chez P. falciparum.
I. Le Paludisme : un fléau mondial.
Le
paludisme est une maladie parasitaire causée par un hématozoaire
du genre
Plasmodium et transmise à l'homme par des moustiques
femelles du genre Anopheles. Il s'agit de la principale maladie
parasitaire humaine. En 1994, le risque de paludisme existait à
des degrés divers dans 100 pays et territoires. Dans 92 pays et
territoires, il y avait transmission du paludisme à Plasmodiumfalciparum
et dans 8 pays, uniquement transmission de P. vivax (Figure
1). Les estimations de la population exposée au risque dans
ces 100 pays et territoires dendémie sont très variables
du fait de la modification de lenvironnement et dune mobilité
accrue de la population. En 1994, on estime que 2,3 milliards de personnes,
cest-à-dire 41% de la population mondiale, vivaient dans des zones
à risque palustre .
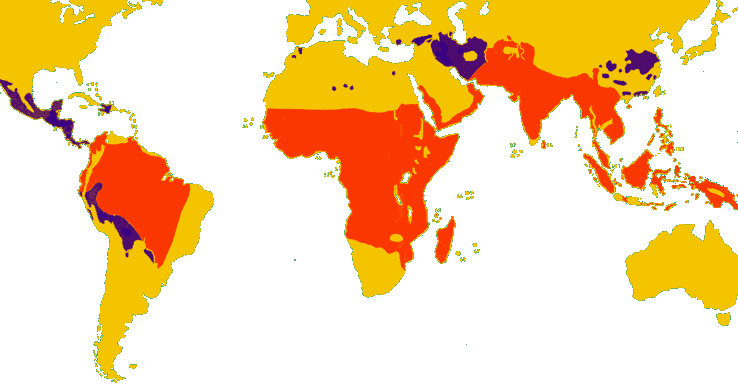
Figure 1 : Evaluation épidémiologique de la situation du paludisme en 1997. En jaune sont figurées les zones dans lesquelles le paludisme a disparu, a été éradiqué ou na jamais sévi ; en violet : zones à transmission limitée ; en orange : les zones où il y a transmission du paludisme.
Des
quatre espèces pathogènes chez l'homme (P. falciparum,
P. vivax, P. malariæ, P. ovale) Plasmodium falciparum
est la plus redoutable. Ce dernier provoque les pathologies les plus sévères
avec complications pulmonaires, rénales, cérébrales
et anémies sévères. Il sévit toute l'année
dans les pays équatoriaux où il peut, cependant, présenter
des recrudescences saisonnières ; dans les régions subtropicales,
il ne survient qu'à la période chaude et humide. La maladie
frappe les enfants en bas âge, les adultes non immuns et les femmes
au cours de leurs premières grossesses. Cest lespèce prédominante
en Afrique tropicale, en Asie orientale, en Océanie et dans la région
amazonienne. Elle est moins fréquente dans le reste du monde. Un
diagnostic précoce et un traitement rapide et approprié de
la maladie sont indispensables pour prévenir la mortalité.
La grande majorité des décès dus au paludisme se produisent
chez les enfants en bas âge de lAfrique tropicale, notamment dans
les régions rurales écartées, mal desservies par les
services de santé. En dehors de lAfrique tropicale, les cas mortels
concernent principalement les sujets non immuns qui arrivent pour la première
fois dans des zones dendémie, comme les ouvriers agricoles, les
travailleurs des mines dor et de pierres précieuses, et les colons
des nouvelles zones de peuplement. Durant les épidémies,
tous les groupes dâge sont touchés. Cest dans les régions
en proie aux conflits armés, au commerce illicite et aux migrations
massives de réfugiés que les problèmes sont parfois
les plus graves.
Comme
nous lavons vu dans le Préambule, actuellement, le paludisme est
en recrudescence dans de nombreux pays. Les moyens de lutte dont on dispose
à lheure actuelle sont sérieusement menacés. Devant
cette situation, il est urgent de mettre en place de nouveaux moyens de
lutte anti-vectorielle ; l'élaboration de nouveaux médicaments
et la mise au point de vaccins sont donc nécessaires. Plusieurs
essais vaccinaux humains ont eu lieu, mais les résultats obtenus
ont été décevants
.
Le vaccin synthétique
SPf66, développé par Patarroyo et son équipe , a fait
lobjet de la plus grande étude jamais mise en place au niveau mondial,
en vaccinologie anti-palustre. Il est apparemment bien toléré
et immunogène chez l'homme , a conféré une protection
partielle lors d'essais dans des zones de faible transmission comme la
Colombie et le Vénézuéla . Un essai mené chez
des enfants en zone dholoendémie (Tanzanie) a donné des
résultats encourageants , qui nont pas été confirmés
par les essais ultérieurs menés en Gambie (Alessandro et
al, 1995), en Thaïlande ou en Tanzanie . Lidentification de nouvelles
cibles vaccinales ou thérapeutiques est, donc, primordiale.
II. Plasmodium : un parasite adapté à son hôte.

Tableau 1 : Quelques espèces des plasmodies, montrant la variété des hôtes intermédiaires.
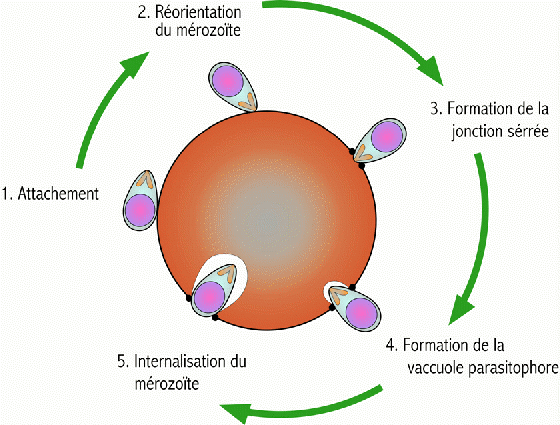
Le
sporozoïte entre alors dans une phase de réplication, au sein
de la vacuole parasitophore, et de prolifération intracellulaire
qui repousse en périphérie le noyau de la cellule et finit
par constituer une masse multinucléée appelée schizonte
qui conduit à la libération de plusieurs dizaines de milliers
de mérozoïtes dans la circulation (Figure
2). Cette phase de multiplication est asymptomatique et dure de
8 à 15 jours, selon les espèces. Contrairement à P.
vivax,
P. falciparum ne possède pas de formes de persistance
hépatique ou hypnozoïtes.
L'apparition des gamétocytes a lieu en général la deuxième semaine qui suit l'infection et ces formes peuvent persister plusieurs semaines après la guérison. A la suite d'une nouvelle piqûre par une Anophèle, les gamétocytes mâles et femelles (au dimorphisme sexuel marqué) sont ingérés avec le repas sanguin.
Il est important de noter que lérythrocyte, ne possédant pas de système de synthèse et de transport des protéines et nexprimant pas de molécules du MHC de classe I ou II à sa surface, est un refuge idéal pour un parasite qui doit perdurer de longues périodes chez son hôte, afin dêtre transmis au moustique.

Figure 2 : Cycle de développement de P. falciparum chez lHomme.
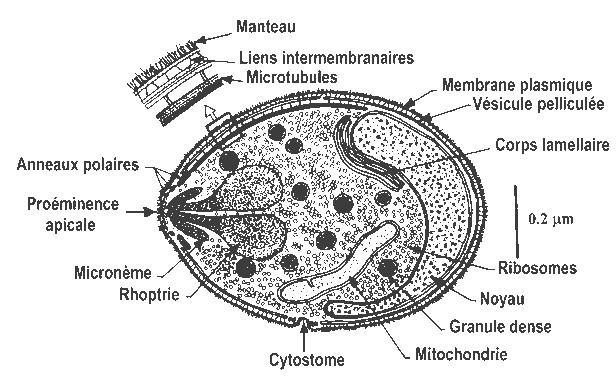
Figure 3 : Représentation schématique de la section dun mérozoïte de Plasmodium falciparum et des ultrastructures de la cellule .
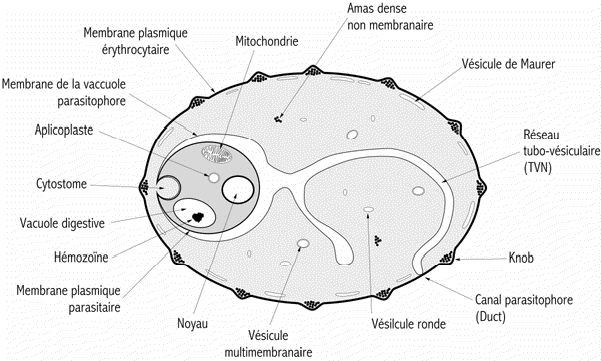
Figure 5: Représentation schématique de lérythrocyte parasité.
Dautre
part, il existe à la surface de la membrane érythrocytaire
des structures semi-cristalines, denses aux électrons, formant des
protubérences, appellées " knobs ". Il est à
noter que P. falciparum est le seul des quatre espèces plasmodiales
humaines à posséder des "knobs" que portent les hématies
infectées au stade trophozoïte âgé et schizonte.
Ces "knobs" contribuent à la séquestration des érythrocytes
parasités au niveau des capillaires périphériques
et dans certains organes comme la rate, les poumons et le cerveau. Ceci
explique que l'on ne détecte pas de parasites au stade schizonte
dans la circulation. Ce phénomène de séquestration
est associé à la gravité des symptômes ; en
effet, l'obstruction des vaisseaux, même partielle, pourrait entraîner
une surpression et une hypoxie locale (effet Sludge).
L'accès
palustre est généralement précédé
de légers symptômes tels que céphalée, fatigue,
courbatures, anorexie, vomissements et diarrhées chez les enfants.
La durée du cycle de P. falciparum étant de 48 heures,
on observe, dans les cas typiques, des pointes de fièvre, tous les
2 jours, contemporaines de la rupture des schizontes, on parle de fièvre
tierce ; plus généralement, quand les parasites ne sont pas
synchronisés, on observe des poussées de fièvre quotidiennes.
L'accès palustre dure de 6 à 10 heures et se décompose,
le plus souvent, en trois phases. Tout d'abord une sensation de froid
intense qui dure une quinzaine de minutes et qui est suivie par une
montée de la température au-dessus de 40°C, hyperpyréxie
qui peut durer plusieurs heures et qui est, éventuellement, accompagnée,
dans les cas graves, de vomissements, de convulsions et de chutes de tension
artérielle. Enfin le malade entre dans une phase de sudation
quand la température redescend. Laccès palustre simple ne
possède aucune manifestation clinique spécifique, doù,
parfois, la difficulté à établir un diagnostique.
Si l'accès palustre présente des complications, on parle
d'accès pernicieux. Ces complications sont: neuropaludisme
avec ou sans coma, anémie normocytaire profonde, insuffisance rénale
aiguë, dème pulmonaire, hypoglycémie, collapsus circulatoire,
hémorragie spontanée avec trouble de la coagulation, convulsions
généralisées et répétées, acidose
métabolique et hémoglobinurie.
Les études épidémiologiques effectuées en zone d'endémie comme celle qui est actuellement menée dans le village de DIELMO (Sénégal) grâce à une coopération entre l'Unité dImmunologie Moléculaire des Parasites, le laboratoire de Parasitologie Médicale (Institut Pasteur), l'Institut Pasteur de Dakar et lIRD ont permis de confirmer, tout en fournissant plus de renseignements, les observations originelles faites depuis longtemps déjà, sur l'existence de deux phases dans le développement de la prémunition . Une première phase qui correspond à l'acquisition d'une immunité clinique, protège les individus contre les symptômes plus ou moins sévères de la maladie. Cette première phase semble peu affecter les niveaux de parasitémie circulante. L'immunité antiparasite, qui se développe dans un deuxième temps, est responsable du contrôle de la parasitémie à un faible niveau.
On
ne connaît que peu de choses sur les mécanismes effecteurs
de la réponse immune chez l'homme, et ceci notamment parce qu'il
est difficile de définir des critères épidémiologiques,
cliniques ou parasitologiques simples susceptibles de caractériser
cet état de prémunition et de définir clairement quels
sont les individus "protégés".
Cependant,
les anticorps semblent jouer un rôle majeur dans l'immunité
contre la phase sanguine. En effet, des expériences de transfert
passif effectuées dans les années soixante et réitérées
plus récemment montrent que les anticorps provenant d'individus
prémunis sont capables de transférer cette protection à
des individus en cours d'accès.
Les
anticorps pourraient agir soit directement en bloquant l'invasion des érythrocytes
par les mérozoïtes, soit en coopération avec les monocytes/macrophages,
par opsonisation puis phagocytose, par mécanismes d'ADCC (Cytotoxicité
Cellulaire Dépendante d'Anticorps) ou mécanismes d'ADCI (Inhibition
Cellulaire Dépendante d'Anticorps) . Par ces mécanismes,
les anticorps contribueraient au contrôle à long terme de
la parasitémie chez les individus prémunis.
La
diversité antigénique de P. falciparum est le reflet
dun polymorphisme allélique important. Plusieurs études
et techniques ont permis de mettre en évidence le polymorphisme
allélique, que ce soit par RFLP ou par typage génétique
disolats de zone dendémie palustre par PCR ainsi que le séquençage
de plusieurs allèles de certains gènes comme MSP-1 . Chez
P.
falciparum, de nombreux antigènes codés par des gènes
en copie unique possèdent un très grand nombre dallèles,
pouvant, parfois, être regroupés en famille alléliques
de séquence primaire très différentes (comme MSP-1
et MSP2
; Figure 6).
Les bases structurelles de ce polymorphisme sont multiples. Il y a, bien sûr, les mutations ponctuelles . Cependant P. falciparum utilise de manière intensive dautres mécanismes afin de générer une telle diversité. Beaucoup dantigènes comportent des séquences répétées ; suivant les allèles, celles-ci peuvent varier en nombre (Antigène S, Pf11.1, GBP 130, CSP), en taille (Antigène S) ou même de cadre de lecture. Enfin la dégénérescence des répétitions assure un degré de diversité supplémentaire (Figure 6).
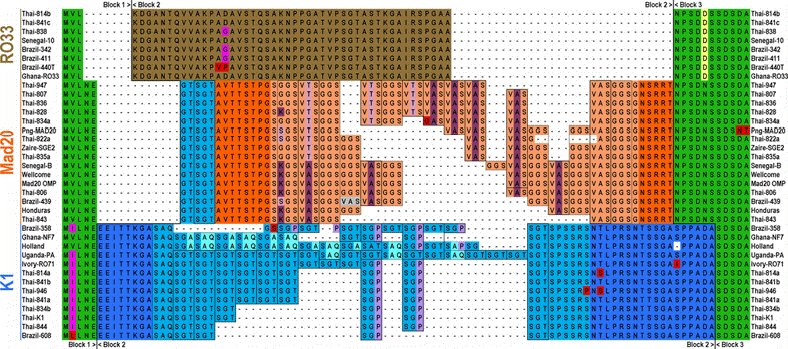
Figure
6 : Alignement des séquences protéiques déduites du
bloc 2 de MSP1.
Les
3 familles allèliques de séquences primaires différentes
sont représentées. Chacune delles varient en fonction du
nombre de répétitions, certaines répétions
sont dégénérées. On peut noter un certain nombre
de délétions, et de mutations ponctuelles (daprès
Miller et alii, 1993 modifié E. Bischoff).
![]()
![]()